Anatomy and Morphology
Plant Structure and Their Functions
Cells
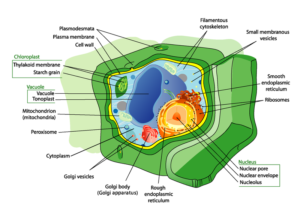 Plants, like animals, consist of millions of individual cells. Both plant and animal cells contain a cell membrane, cytoplasm, nucleus, and other organelles. Plant cells differ from animal cells in that they have a stiff cell wall and a large vacuole for storing water and other nutrients. Cell walls consist of cellulose, a type of carbohydrate that helps the plant maintain its structure. Some cells, especially those found in leaves and stems, can contain chloroplasts which are organelles that contain cylindrical stacks called grana. In these grana is a pigment called chlorophyll that gives plants a green color and allows them to capture sunlight and turn it into sugar (energy) for the plant.
Plants, like animals, consist of millions of individual cells. Both plant and animal cells contain a cell membrane, cytoplasm, nucleus, and other organelles. Plant cells differ from animal cells in that they have a stiff cell wall and a large vacuole for storing water and other nutrients. Cell walls consist of cellulose, a type of carbohydrate that helps the plant maintain its structure. Some cells, especially those found in leaves and stems, can contain chloroplasts which are organelles that contain cylindrical stacks called grana. In these grana is a pigment called chlorophyll that gives plants a green color and allows them to capture sunlight and turn it into sugar (energy) for the plant.
Groups of cells with similar functions are called tissues. These tissues group together to form the various parts of a plant, including roots, stems, buds, leaves, flowers, and fruits.
Roots
Roots typically develop from the lower portion of a plant or cutting. The principal functions of roots are to absorb nutrients and water, anchor the plant in the soil or other growing medium, to provide physical support for the stem, and serve as food storage organs. In some plants, roots are used as a means of propagation—either naturally or through human intervention.
The structure and growth habits of roots have a pronounced effect on a plant’s size and vigor, method of propagation, adaptation to certain soil types, and response to cultural practices and irrigation. Roots of certain vegetable crops are also important food sources.
Root Development
A root has three major developmental zones.
- The zone of cell division includes the root cap, which protects the interior cells as the root pushes through the soil, and the root apical meristem, an area of division or growth. The root apical meristem produces new cells that contribute to the root cap as well as to the root itself.
- The second zone, behind the meristem, is called the zone of elongation. Here cells take in food and absorb water, thereby increasing in size and pushing the tip of the root through the soil.
- The third major root zone is the maturation zone, where the majority of absorption occurs and cells undergo changes to become specific tissues, such as epidermis, cortex, or vascular tissue.
- The epidermis is the outermost layer of cells surrounding the root. These cells are responsible for absorbing water and minerals dissolved in water.
- The cortex outermost layer of the stem or root of a plant, bounded on the outside by the epidermis and on the inside by the endodermis.
- The Vascular tissue is located in the center of the root and conducts food and water.
Root hairs begin to emerge in the zone of maturation, developing as outgrowths of the epidermal cells. The majority of the water and nutrients absorbed by the plant enter through the root hairs. Most plants produce root hairs that live only a few days or a few weeks. New root hairs form as roots continue to grow through the soil. Older portions of a root do not have root hairs and cannot absorb water.
|
Root Systems and Adaptations
Plants generally have one of two types of root systems: taproot or fibrous root. At the end of the embryo of a seedling, the tip that will develop into a root is called a radicle. A taproot forms when the primary root continues to elongate downward into the soil. The taproot becomes the central and most important feature of the root system. Taproots have a somewhat limited amount of secondary (lateral or side) branching. Some trees, such as pecan trees, have a long taproot with very few lateral or finer, fibrous roots. This makes these trees difficult to transplant and necessitates planting in deep, welldrained soil. The taproots of carrots, parsnips, and salsify are the main edible parts of these crops.
If plants that normally develop a taproot are undercut so that the taproot is severed early in the plant’s life, the root will lose its taproot characteristic and develop a fibrous root system. Undercutting is done intentionally in commercial nurseries to force trees to develop a compact, fibrous root system, which contributes to a higher rate of transplanting success.
A fibrous root system is one in which numerous lateral roots develop. These roots branch repeatedly and form the plant’s feeding root system.
Fleshy roots become food reservoirs that store surplus food for the winter or other adverse periods to be used by the plant when it is able to renew its growth. Carrots, turnips, and beets have primary or taproots containing food. Sweet potatoes and dahlia tubers are secondary roots transformed into tuberous roots packed with food.
Aerial roots form freely on some land and water plants in a favorable, moist environment. These aerial roots enable climbers such as philodendrons, orchids, and air plants to attach themselves to a structure for anchorage and support. Aerial roots absorb water from the air. Many aerial roots are also adventitious, meaning they arise from stem tissue. They are often fleshy or semi-fleshy and function as reservoirs for water storage.
Factors That Influence Root Growth
- Roots grow best in soil with an adequate balance of moisture and air.
- Soil fertility and pH determine the rate and depth of root growth.
- Lack of oxygen in the soil limits growth, leading to a decrease in overall water and nutrient absorption.
- Soil temperature will limit root growth. Roots grow best in the soil if the temperature is between 75-95.
- Soil compaction limits root growth.
Stems
Stems are structures that support buds, flowers, and leaves. The major internal part of a stem is its vascular tissue system, which includes xylem, phloem, and in most plants, the vascular cambium. The vascular tissue system transports food, water, and minerals and provides support for the plant. The conducting elements in the xylem transport water and minerals, while those elements in the phloem conduct food. Both xylem and phloem can conduct hormones.
The two classes of flowering plants, monocots, and dicots, both contain xylem and phloem, but they are arranged in different patterns. In a monocot stem, the xylem and phloem are paired into discrete vascular bundles; these bundles are scattered throughout the stem like a bundle of straws.
In a dicot stem, the vascular system develops in a ring pattern. The ring of phloem is near the bark or external covering of the stem and becomes a component of bark in mature stems. In dicot woody plants, just inside the phloem is the vascular cambium, a site of cell division and active growth. The vascular cambium is responsible for a stem’s increase in diameter as it produces both secondary phloem and xylem tissues. Xylem tissue forms a ring just inside the vascular cambium, thereby forming the sapwood and heartwood.
Woody Stem Parts and Their Functions
|
Understanding the difference between monocot and dicot vascular systems is important to gardeners for several reasons:
- A dicot tree can die if a string trimmer girdles the tree’s trunk. This can occur while someone trims the surrounding grass with a Weed Eater or other string trimmer. Such a wound can sever the vascular system just inside the bark. The same treatment of a
monocot palm tree does minimal damage because the vascular system is distributed throughout the entire trunk. - Dicots have multiple meristems, so cutting off a tip just redirects growth. Monocots, however, have only one apical meristem, so cutting the top off a palm kills the tree.
- Monocots do not have secondary meristem cells, so they grow taller but not wider. Dicots with secondary meristem cells grow wider as well as taller.
- Some selective herbicides kill only dicots.
- Other selective herbicides kill only monocots.
The stem area where leaves are attached (or were once attached) is called a node. Within the node is a bud, which is an area of great cellular activity and growth. The area between nodes is called an internode. The length of an internode depends on many factors. Low fertility, for instance, decreases internode length. Too little light causes a long internode and a spindly stem, an occurrence known as stretching or etiolation. Growth produced early in the season often results in the greatest internode length; internode length decreases as the growing season nears its end. Vigorously growing plants tend to have greater internode lengths than unhealthy plants. Internode length also varies with competition from surrounding stems or developing fruit. If the energy for a stem has to be divided between three or four stems, or if the energy is diverted into fruit growth, the internode length will be shorter.
Stems can be long, with great distances between leaves, buds, and branching, or compressed, with short distances between leaves, buds or branching. The distance between leaf nodes is a good indication of how fast the tree will grow.
Types of Stems
A shoot is a general term used to describe a young stem with leaves. A twig is a woody stem in the current or previous year’s growth. A branch is a stem that is more than a year old and may have lateral stems. A trunk is the main stem of a woody plant. Woody stems contain relatively large amounts of xylem tissue in the central core and are typical of most fruit trees, flowering trees, and shrubs.
A cane is a stem with a relatively large pith (soft spongy tissue in the center of a stem) and a life expectancy of only one or two years. Examples of plants with canes include roses, grapes, blackberries, and raspberries. Herbaceous stems contain only small amounts of xylem tissue and usually live for one growing season.
Modified Stems
When we think of a stem, we tend to picture typical trunks and branches. Modified stems can be grouped into two categories: above ground and below ground. Above ground modified stems are crowns, stolons, and spurs. Below ground stems are bulbs, corms, rhizomes, and tubers.
Above ground Stems
- Crowns (on strawberries, dandelions, and African violets) are compressed stems with leaves and flowers on short internodes.
- A stolon is a specialized stem that grows on the soil surface and forms a daughter plantlet at one or more of its nodes. Some plants that have stolons, such as periwinkle Vinca minor, make good ground covers because they spread easily. Stolons are sometimes called runners, but technically runners are a subgroup of stolons. Examples of plants with runners are strawberries and spider plants.
- Spurs are short, stubby, lateral stems that arise from the main stem and are common on such fruit trees as pears, apples, and cherries. Spurs are where these trees can bear fruit. If severe pruning is done close to fruitbearing spurs, the spurs can revert to long, nonfruiting stems.
Underground Stems
Many types of modified stems can be found beneath the soil. These belowground stems are often confused with roots. If you find the presence of buds, leaf scars, or nodes, you have an underground stem, not a root. These modified stems are energy-storage organs.
- Tubers are fleshy modified stems with nodes that produce buds. A potato is a tuber. The eyes of a potato are actually the nodes on the stem. Each eye contains a cluster of buds. A small piece of potato, as long as it contains an eye, can produce an entirely new potato plant. Some plants, such as begonias and cyclamens, produce an enlarged, modified underground tuberous stem that is short and flat. Unlike true tubers that have buds scattered on all sides, tuberous stems have shoots only on their tops.
- Rhizomes are specialized stems that grow horizontally at or just below the soil surface and serve as a storage organ and a means of propagation in some plants. Aerial stems with leaves grow from one end of the rhizome, and roots grow from the other end. Some rhizomes are compressed and fleshy, such as those of irises. They can also be slender with elongated internodes as in bent grass. Johnson grass (Sorghum halepense) is a hated weed because of the spreading capability of its rhizomes.
- Bulbs are shortened, compressed, underground stems surrounded by thickened, fleshy leaves that envelop a central bud located at the tip of the stem. Tulips, lilies, daffodils, and onions are plants that produce bulbs. If you cut through the center of a tulip or daffodil bulb in November, you can see all the flower parts in miniature within the bulb.
- Corms can have shapes similar to bulbs but do not contain fleshy scales. A corm is a solid, swollen stem with scales that are covered with reduced, scale-like leaves. The new stem grows from a bud on top of the corm. Some plants produce a modified stem that is referred to as a tuberous stem. Examples are crocus, tuberous begonia, and cyclamen. The stem is shortened, flattened, enlarged, and underground. Buds and shoots arise from the crown; fibrous roots occur on the bottom of the tuberous stem.
Thorns, spines, and prickles are terms you may see used interchangeably to describe the sharp projections on plants that protect them from being eaten by herbaceous animals. There are, however, botanical differences between these projections. Thorns are a modification of a stem or branch, which means they can be branched or not, have leaves or not, and they arise from a bud. A black locust tree (Robinia pseudoacacia) has thorns. Spines are modified leaves, leaflets, petioles, or stipules. Blackberries and wintergreen barberry (Berberis juliane) have spines. Despite the adage, every rose does not have thorns. The sharp projections on roses are modified epidural cells called prickles. Prickles can occur anywhere along the plant and are much easier to remove than thorns or spines.
Buds
A bud is an undeveloped shoot from which new leaves or flower parts arise. Buds of trees and shrubs of the temperate zone typically develop a protective outer layer of small, leathery bud scales. Annual plants and herbaceous perennials have naked buds in which the outer leaves are green and somewhat succulent. Buds are named by their location on the stem surface. Terminal buds are those located at the end of a stem. Lateral buds occur on the sides of the stem. Most lateral buds grow in the axil of a leaf (the small angle between the plant’s stem and the leaf attachment) and are called auxiliary buds. In some instances, more than one bud is formed. Adventitious buds are those that arise at sites other than in the terminal or auxiliary positions. Adventitious buds may develop from the internode of the stem, at the edge of a leaf blade, from callus tissue at the cut end of a stem or root, or laterally—from a plant’s roots.
A leaf bud consists of a short stem with immature leaves. Such buds develop into leafy shoots. Leaf buds are often more pointed and less plump than flower buds.
A flower bud consists of a short stem with embryonic floral parts.
Leaves
The primary function of most leaves is to absorb sunlight for the production of plant sugars in a process called photosynthesis. Each leaf develops as a flattened surface that provides a large area for efficient absorption of light energy. The leaf is supported away from the stem by a stem-like appendage called a petiole. The base of the petiole is attached to the stem at the node.
Parts of a Leaf
The blade of a leaf is the expanded, thin structure on either side of the midrib. The blade is usually the largest and most conspicuous part of a leaf. The petiole varies in length and can be lacking entirely (described as sessile or stalkless). Stipules are leaflike structures that often occur in pairs and are sometimes fused. Stipules are usually found at the base of the petiole or on the node of a stem (as on, willow, strawberry, and geranium) or occasionally fused to the petiole (as on rose, crabapple, and Callery pear). Stipules often drop as leaves mature, but those that persist or enlarge are good identification features.
The leaf blade consists of several tissue layers. The epidermis is the complex tissue layer that covers a leaf’s upper and lower surfaces and protects internal tissues. How the cells in the epidermis are arranged determines the texture of the leaf surface. Some leaves have hairs, called trichomes, that are extensions of certain cells of the epidermis. African violets have so many hairs that the leaves feel like velvet, while other species may have very few to no trichomes.
The cuticle is a thin layer covering the epidermis and consists of waxes and a substance called cutin. This substance protects the leaf from dehydration and prevents penetration of some disease-causing organisms. The amount of cuticle material produced by epidermal cells correlates to the intensity of sunlight to which leaves are exposed. Plants grown in the shade should be moved into full sunlight gradually to allow the cutin layer to build and protect leaves from the shock of rapid water loss or sunscald. The cuticle also repels water and can shed pesticides if surfactants or soaps are not used as a sticking agent. This is why many pesticide manufacturers include a spray additive that adheres to or penetrates the cuticle.
A leaf’s epidermal layer also contains small openings, each of which is called a stoma. The plural for stoma is stomata. Each stoma is surrounded by two guard cells. A stoma and its guard cells are called a stomatal complex. The bending of the two guard cells regulates the passage of water, oxygen, and carbon dioxide into or out of the leaf. The opening and closing of the stomata by their guard cells are determined by the different environmental conditions to which the leaves are exposed. Conditions that would cause a higher rate of water loss from plants (high temperature, low humidity) stimulate guard cells to close. In mild weather, stomata often remain open. With very few exceptions, guard cells usually close the stomata in the absence of light.
Leaves as a Means of Identifying Plants
Leaves, especially the shapes of their blades and margins (edges), are useful in identifying species and cultivars of horticultural plants. Simple leaves are those in which the leaf blade is a single continuous unit. A compound leaf consists of a blade that is divided into two or more similar structures called leaflets. Although compound leaves may resemble a branch stem with several leaf blades attached, leaflets do not have an axillary bud at their base. An axillary bud demarcates the true base of the entire leaf organ structure.
Most compound leaves are pinnately compound, which means that the leaf is structured like a feather, with all leaflets arranged on both sides of a central axis, called the rachis. Common examples are leaves of ashes, pecans, and Chinese pistache. Some leaves may be doubly (bipinnate) or triply (tripinnate) compounds, meaning the leaflets are further divided. Honey locust and nandina (which is triply compound) are examples. Palmately compound leaves are less common than pinnately compounds. Palmately compound leaves resemble a hand, with all leaflets emerging from the end of the petiole. Examples include buckeye, chaste tree, horse chestnut, and Schefflera species.
Some blades are interrupted along the margin and can be mistakenly identified as compound leaves. Lobed leaves, for example, are simple leaves with indentations reaching halfway or less to the center of the leaf, as in a pin oak. Blades with deep indentations reaching close to the midrib yet that are connected by narrow bands of blade tissue (such as ferns) are simple leaves described as pinnatifid.
Leaf Morphology as a Means of Identification
The number of taxonomic terms can be overwhelming at first, but familiarizing yourself with them will help with plant identification. Plant identification is the first step in diagnosing any plant problem. Careful observation of plants can help us recognize the differences and similarities among plants and plant families.
Leaf Blade
These are some of the most common leaf shapes:
- Linear—Narrow, five or more times longer than wide; approximately the same width throughout
- Lanceolate—Lance-shaped; broadest below the middle and tapering toward the apex
- Elliptical—Ellipse-shaped, broadest in the middle, tapering to an acute or rounded apex and base; two or three times longer than wide
- Ovate—Egg-shaped, widest below the middle, and tapering toward the apex; two to three times longer than wide
- Cordate—Heart-shaped or broadly ovate; tapering to an acute apex, with the base turning in and forming a notch where the petiole is attached.
Apexes and Bases
These are some of the most common apex and base shapes for leaves:
- Acuminate—A prolonged apex tapering to a long, narrow point
- Acute—Ending in an acute angle, with a sharp point
- Obtuse—Ending in an obtuse angle
- Retuse—A rounded apex with a shallow notch
- Emarginate—A deep notch
- Sagittate—Arrowhead-shaped, with two pointed lower lobes projected downward
- Truncate—Having a relatively square end
- Attenuate—A prolonged base with thin tissue extending along part of the petiole
- Cuneate—Wedge-shaped, tapering at an acute angle toward the base
Leaf Margins
Leaf margins are especially useful in the identification of some plants:
- Entire—A smooth edge with no teeth or notches
- Sinuate—Having a pronounced wavy margin
- Crenate—Having rounded teeth
- Dentate—Having large blunt teeth ending in an acute angle, pointing outward
- Serrate—Having small, sharp teeth pointing toward the apex
- Incised—Margin cut nearly to the midrib into sharp, deep, irregular incisions
- Lobed—Incisions extend less than halfway to the midrib
- Cleft—Incisions extend more than halfway to the midrib
Leaf Textures
Leaf texture is another way to identify a plant. These are some common leaf textures:
- Succulent—Fleshy, soft, and thickened in texture; modified for water storage
- Coriaceous—Leather-like, tough
- Glabrous—Smooth, shiny, hairless surface
- Downy—Covered with very short, weak, and soft hairs
- Pubescent—A hairy surface
- Tomentose—Covered with matted, wooly hairs
- Hirsute—Pubescent with coarse, stiff hairs
- Hispid —Rough with bristles, stiff hairs, or minute prickles
- Scabrous—Rough to the touch, with the texture of sandpaper
Leaf Arrangement Along a Stem
Leaves are attached to stems in various patterns, and these patterns can also be used to identify plants. Alternate leaves are arranged along the stem with only one leaf at each node, alternating on each side of the stem. This arrangement is most common. Opposite leaves occur when two leaves at each node are located across from one another. A whorled arrangement is rare and requires three or more leaves at a node. Plants exhibiting whorled leaves may have opposite leaves as well, such as glossy abelia (Abelia × grandiflora) and cape jasmine (Gardenia augusta G. jasminodies).
Venation
Plants vary with respect to the venation pattern in multiple organs but nowhere are these differences more noticeable than in leaves. Leaves with linear venation are those with numerous lateral veins that run essentially parallel to each other and are connected laterally by minute, straight veinlets. A common type of parallel venation is that found in plants of the grass family, where the veins run parallel from the base to the apex of the leaf. Another type of parallel venation occurs in plants such as bananas, cannas, and pickerelweed. In these plants, the parallel veins run through the center then turn laterally to the margin. Leaves with parallel venation occur in monocot plants. They do not have a midrib but may have one to several larger veins with smaller parallel veins in between them.
Leaves with netted venation have secondary veins that branch from a conspicuous midrib and then subdivide into finer veinlets, which unite into a complicated network. This system of enmeshed veins gives the leaf more resistance to tearing than most parallel-veined leaves. Netted venation may be either pinnate or palmate. In pinnate venation, the secondary veins extend laterally from the midrib to the edge, as in apple, cherry, and peach trees. Palmate venation occurs in grapevine and maple leaves, where several principal veins extend outward, like the ribs of a fan, from the petiole near the base of the leaf blade. Generally, leaves with netted venation occur on dicots.
Several distinct types of leaves occur on plants. Scale leaves or cataphylls are found on rhizomes and are also the small, leathery leaves that enclose and protect buds. Cotyledons are modified seed leaves that are part of the seed embryo and early seedling. Cotyledons typically serve as energy storage structures for germination and establishment of a seedling’s first true leaves. Other specialized leaves include sharp spines that protect the plant and tendrils that assist in supporting stems. Storage leaves are found in bulbous plants and succulents. Bracts are specialized leaves that may be brightly colored. The showy structures on dogwoods and poinsettias are not petals. They are bracts, modified to attract pollinators.
Flowers
Although we appreciate flowers for their beauty and scent, a flower’s sole function is sexual reproduction Plants like grasses and pines that are wind pollinated have no need to attract an insect or animal and so expend little energy on beauty but instead produce copious amounts of pollen for the wind to disperse. Other plants, however, have developed fragrant, pale, night-blooming flowers to attract pollinator moths.
Most fragrant, showy flowers with nectaries have evolved to attract insect pollinators to ensure the reproductive success and continuance of the plant species. Markings on petals signal to insects how to approach and where to land. Bee-pollinated flowers reflect ultraviolet light that is visible to insects but not humans. Some male insects are tricked into transferring pollen by flowers that look and smell like females of an insect species. Fragrant flowers attract bees, butterflies, moths, and other insects. Flowers with foul odors attract flies, beetles, and similar insects.
Other animals, such as birds and bats, also pollinate flowers. Some flowers have landing platforms or nectaries at the end of long tubes to match the body parts of the animals that perform pollination.
Parts of the Flower
As the reproductive structure of the plant, the flower contains pollen-producing structures, ovule-bearing structures, and sterile structures such as petals, sepals, and nectaries.
A peduncle is a stem that supports the flower or flowers. Sepals are small, usually green, leaf-like structures on the base of the flower. Sepals collectively are called the calyx. Petals are usually the highly colored portions of the flower. They may contain specialized structures that produce molecules associated with a scent called osmophores, or they may produce nectar in structures known as nectaries. The petals collectively are the corolla. The number of petals and sepals on a flower, and the form they take, are often used to identify plant families and genera. The calyx and corolla together are called the perianth.
The receptacle is the expanded tip of a flower stalk that bears the flower’s sexual organs. The pistil, located in the center, is the female structure of a flower. Some plants have flowers with no pistil; others have one or more pistils. The totality of the pistils is referred to as a gynoecium (“house of woman”). A single pistil consists of a stigma, style, and ovary. The stigma is located at the apex and is the site of pollination. The style is the stalk between the stigma and the ovary. The ovary is the swollen part at the base of the style; it contains ovules, each of which produces a single egg cell. After the egg is fertilized, the ovule develops into a seed, and the ovary matures into the fruit.
The stamen is the male reproductive organ. Each stamen consists of an anther, containing one to several pollen-producing chambers, and a long, supporting stalk called the filament. This filament holds the anther in position so that pollen can be disbursed by wind or carried to the stigma by insects, birds, or other pollinators.
Types of Flowers
A flower that has sepals, petals, stamens, and pistils is called a complete flower. Incomplete flowers lack one or more of these parts. Grass plants, for example, typically do not have sepals or petals. If a flower contains both male and female reproductive parts, it is said to be perfect. Flowers that lack one of the reproductive structures, either the stamens or pistils, are known as imperfect flowers. An imperfect flower is also, therefore, by definition, incomplete.
Some plants bear only “male” flowers (staminate) or only “female” flowers (pistillate). Species in which the sexes are separated into staminate and pistillate plants are called dioecious (two houses). Most hollies are dioecious. Therefore, to obtain holly berries, we must plant female plants with a male plant nearby. Plants that have separate male and female flowers on the same plant are monecious (one house). Corn, watermelon, and pecan trees are examples of monecious plants. Some monecious plants, such as cucumbers and squash, bear only male flowers at the beginning of the blooming season but later develop flowers of both sexes.
Types of Inflorescences
Some plants bear only one flower per stem or branch; these are called solitary flowers. Other plants produce an inflorescence, a term that refers to a cluster of flowers and how they are arranged on a floral stem. Most inflorescences can be classified into two groups, indeterminate and determinate. Indeterminate inflorescences have an axis terminating in meristematic tissue, which continues to develop until the inflorescence is complete. Flowers bloom in sequence from the bottom to the top of the axis or inward, starting from the outside. There are five basic types of indeterminate inflorescences. A raceme has an elongated axis with stalked flowers. A spike has an elongated axis with sessile (stalkless) flowers. A corymb has a flat-topped or hemispherical raceme with floral stalks of varying lengths arising up the axis. An umbel is similar, but the branches arise from one point at the top of the axis. A panicle is a branched raceme.
Determinate inflorescences have a flower that terminates the axis early in its development. Flowers bloom in sequence down from the tip of the axis or progressively outward from the middle. A head has all flowers attached to a disk; examples are sunflowers and Echinacea species. A cyme is a broad, flat-topped, or hemispherical inflorescence with flower clusters of threes, in which the floral stalks are similar in length. Some inflorescences can have an indeterminate central axis and determinate lateral branches, such as buckeyes.
Day Length and Flowering
Light duration, or photoperiod, refers to the amount of time that a plant is exposed to sunlight. The ability of many plants to flower is controlled by photoperiod. When scientists first recognized the concept of a photoperiod, they believed that the duration of exposure to light triggered flowering. Later they discovered that it is not the light but the uninterrupted dark that triggers floral development. Plants can be classified into three categories, depending upon their flowering response to the duration of darkness: short day, long day, or day neutral.
Short-day plants produce flowers only when the period of darkness exceeds a critical minimum called the critical night length. Short-day plants include many spring- and fall-flowering plants such as chrysanthemum and poinsettia. Long-day plants form flowers only when the period of darkness is less than the critical night length. They include almost all of the summer-flowering plants, such as black-eyed Susan (Rudbeckia fulgida) and California poppies (Eschscholzia californica) , as well as many vegetables, including beets, radishes, lettuce, spinach, and potatoes.
Because chrysanthemums flower under the short-day conditions of spring or fall, the method for manipulating the plant into experiencing short days is simple. If long days are predominant, a shade cloth can be drawn over the chrysanthemum for 13 hours daily to block out light until flower buds are initiated. To bring a long-day plant into flower when sunlight is not present longer than a critical minimum, artificial light can be added until flower buds initiate.
Day-neutral plants produce flowers regardless of day length. Some plants do not really fit into any category but can respond to combinations of day lengths. Petunias will flower regardless of day length but will do so earlier and more profusely under long periods of daylight exposure (short periods of darkness).
Fruit
Fruits are important to human nutrition and often have a prominent place in a garden. Fruit is a ripened ovary that contains one or more seeds. The fruit functions as protection for the seed until it is ready for dispersal. Many fruits are attractive to animals, thus a fruit’s seed can be eaten, pass through a digestive tract, and end up far away from the parent plant, spreading genes and ensuring the long-term survival of the plant. Some fruits rely on wind to disperse the seeds like the fluff on a dandelion or the winged samara of a maple.
Types of Fruit
Fruits can be simple or compound. Simple fruits are formed in a flower with one ovary or several fused ovaries. Cherries, nectarines, peaches, and plums are drupes with a single hard seed in the center. Apples, pears, and quince are known as pomes. Each is a single fruit, but inside it has five fused ovaries, hence the five-pointed star shape when it is cut around its equator. Cantaloupes, cucumbers, eggplant, pumpkins, tomatoes, and watermelons are considered berries; they have several seeds inside a soft outer shell. Some fruits are dry and do not have the typical flesh that comes to mind when one hears the word “fruit.” Dry fruits can be either dehiscent and open when ripe or indehiscent and remain closed when ripe. Milkweed, poppies, and legumes are examples of dehiscent fruits. Indehiscent fruits remain closed when ripe and depend on some other mechanism, such as decay or predators, to release their contents. Indehiscent fruits include pecans, peanuts, or maples.
Compound fruits are either aggregate or multiple. If a single flower has several ovaries that stay together, it forms an aggregate fruit. Raspberries are aggregate fruits, technically aggregate drupes. If several flowers are tightly clustered together on one structure and these ovaries remain close together as the fruit ripens, they produce multiple fruits. Pineapples and figs are examples of multiple fruits.
Seeds
Seeds can be a flower’s end product. A seed contains three parts:
- A miniature plant (embryo) that is in a state of dormancy.
- An endosperm, which is a stored food source for the embryo that can include proteins, carbohydrates, or fats.
- A seed coat or layer around the seed protects the embryo from diseases, insects, and moisture until it is time for germination.
Germination occurs when the proper temperature, moisture, oxygen, and light conditions are met. As water is drawn in through the seed coat, the radicle is the first part of the seed to appear. The radicle will develop into the primary root and will eventually sprout root hairs and lateral roots. The hypocotyl will eventually become the stem. The first leaves or cotyledons are usually shaped differently than the leaves the mature plant will produce. Those seeds with two cotyledons are dicots and those with one are monocots.